蚯蚓
說到蚯蚓,不管是大人小孩應該都不陌生。然而一般人對於蚯蚓的印象,不外乎是體長 10 公分左右,可以用來釣魚;有些人可能還曾見過身長超過 20 公分宛如小蛇的蛇蚯蚓;另外,許多人還知道蚯蚓可以翻動泥土,促進土壤中養分的循環,有利於植物的生長。可是,除此之外,人們對於蚯蚓的瞭解,卻十分有限。
蚯蚓(earthworms)是大型陸棲寡毛類的俗稱,但亦包含少數水生的物種 ,屬於環節動物門(Annelida)的環帶綱(Clitellata),主要分布於土壤表層。 另有一些小型的陸棲寡毛類,俗稱線蚓(potworms),身體呈白色,體長小於 2 公分。 此外,俗稱「紅蟲」的水生絲蚯蚓,主要分布在各種淡水水域,一般體型較小,以水中的有機物質為食物來源。許多人以為蚯蚓的種類很少,不是小隻的普通蚯蚓,就是大隻的蛇蚯蚓,實則不然,以台大校園為例,目前記錄的就有 11 種蚯蚓,而目前台灣確定學名的蚯蚓,更有 73 種之多,再加上許多未定名與未發現的種類,台灣的蚯蚓種數應該不只如此。
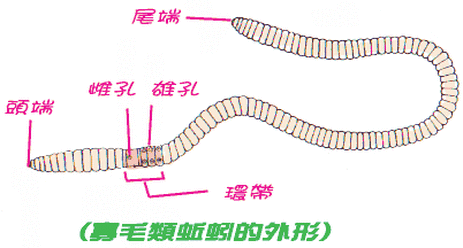
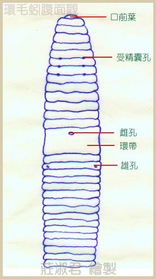
環節動物門,顧名思義,就是身體呈環狀分節,一般而言,蚯蚓的分節多在 80節以上。外觀上除了分節之外,成熟的蚯蚓在靠近頭部的地方,有體節會瘉合成環帶。以環毛蚓為例,環帶上有雌孔,是蚯蚓排出卵子的地方。在環帶的後方則有兩個雄孔,是蚯蚓排出精子的地方;雄孔四周常有乳頭狀突起,稱為乳突,雄孔與乳突是分辨蚯蚓種類的重要依據。然而,大多數的蚯蚓僅由外觀是無法判定其種類的,需要解剖,以其內部構造如受精囊、攝護腺及腸盲囊等的數量、大小、形狀與位置來鑑定其種類。
說到蚯蚓,不管是大人小孩應該都不陌生。然而一般人對於蚯蚓的印象,不外乎是體長 10 公分左右,可以用來釣魚;有些人可能還曾見過身長超過 20 公分宛如小蛇的蛇蚯蚓;另外,許多人還知道蚯蚓可以翻動泥土,促進土壤中養分的循環,有利於植物的生長。可是,除此之外,人們對於蚯蚓的瞭解,卻十分有限。
蚯蚓(earthworms)是大型陸棲寡毛類的俗稱,但亦包含少數水生的物種 ,屬於環節動物門(Annelida)的環帶綱(Clitellata),主要分布於土壤表層。 另有一些小型的陸棲寡毛類,俗稱線蚓(potworms),身體呈白色,體長小於 2 公分。 此外,俗稱「紅蟲」的水生絲蚯蚓,主要分布在各種淡水水域,一般體型較小,以水中的有機物質為食物來源。許多人以為蚯蚓的種類很少,不是小隻的普通蚯蚓,就是大隻的蛇蚯蚓,實則不然,以台大校園為例,目前記錄的就有 11 種蚯蚓,而目前台灣確定學名的蚯蚓,更有 73 種之多,再加上許多未定名與未發現的種類,台灣的蚯蚓種數應該不只如此。
環節動物門,顧名思義,就是身體呈環狀分節,一般而言,蚯蚓的分節多在 80節以上。外觀上除了分節之外,成熟的蚯蚓在靠近頭部的地方,有體節會瘉合成環帶。以環毛蚓為例,環帶上有雌孔,是蚯蚓排出卵子的地方。在環帶的後方則有兩個雄孔,是蚯蚓排出精子的地方;雄孔四周常有乳頭狀突起,稱為乳突,雄孔與乳突是分辨蚯蚓種類的重要依據。然而,大多數的蚯蚓僅由外觀是無法判定其種類的,需要解剖,以其內部構造如受精囊、攝護腺及腸盲囊等的數量、大小、形狀與位置來鑑定其種類。

界: 動物界 Animalia
門: 環節動物門 Annelida
綱: 環帶綱 Clitellata
亞綱:寡毛亞綱 Oligochaeta
目: 單向蚓目 Haplotaxida
包氏重胃蚓 (Dichogaster bolaui )
門: 環節動物門 Annelida
綱: 環帶綱 Clitellata
亞綱:寡毛亞綱 Oligochaeta
目: 單向蚓目 Haplotaxida
包氏重胃蚓 (Dichogaster bolaui )
生殖
蚯蚓是雌雄同體,雖然有越來越多有關蚯蚓可行孤雌生殖,但蚯蚓一般描述仍多稱它們為是雌雄同體異體受精。大部分的蚯蚓交配是在地底,會爬出地表交配的蚯蚓種類只佔全部蚯蚓的少部分,因此觀察並不容易。在國外相關報告中, Lumbricus terrestris (普通蚯蚓)是以相互受精 (cross-fertilization)的方式進行交配 (Edward and Bohlen, 1996):兩隻對向爬行蚯蚓接觸後,若願進行交配,會將自己的雄孔對準對方的受精囊孔,把精子射入對方的受精囊中,若受精囊孔數多於雄孔數時,雄孔會先對準最後一對(最遠離頭部的一對)受精囊孔,將精子送入受精囊內後,再向後縮使雄孔對到前一對受精囊中,依此類推,最後互相離開。受精後的蚯蚓,會由雌性生殖孔(雌孔)產生卵,並由環帶處分泌黏液蛋白包住卵,隨著肌肉的推進至受精囊孔處,釋放精子,完成受精,最後黏液蛋白硬化由頭部推出形成卵繭,每個卵繭中可以含有一至多個受精卵,不同種類孵化時間不一,幼蚓孵化後會自行鑽出卵繭生活。
蚯蚓是雌雄同體,雖然有越來越多有關蚯蚓可行孤雌生殖,但蚯蚓一般描述仍多稱它們為是雌雄同體異體受精。大部分的蚯蚓交配是在地底,會爬出地表交配的蚯蚓種類只佔全部蚯蚓的少部分,因此觀察並不容易。在國外相關報告中, Lumbricus terrestris (普通蚯蚓)是以相互受精 (cross-fertilization)的方式進行交配 (Edward and Bohlen, 1996):兩隻對向爬行蚯蚓接觸後,若願進行交配,會將自己的雄孔對準對方的受精囊孔,把精子射入對方的受精囊中,若受精囊孔數多於雄孔數時,雄孔會先對準最後一對(最遠離頭部的一對)受精囊孔,將精子送入受精囊內後,再向後縮使雄孔對到前一對受精囊中,依此類推,最後互相離開。受精後的蚯蚓,會由雌性生殖孔(雌孔)產生卵,並由環帶處分泌黏液蛋白包住卵,隨著肌肉的推進至受精囊孔處,釋放精子,完成受精,最後黏液蛋白硬化由頭部推出形成卵繭,每個卵繭中可以含有一至多個受精卵,不同種類孵化時間不一,幼蚓孵化後會自行鑽出卵繭生活。

紅蚯蚓(Eisenia sp.)的交配行為,是在土壤中進行,台灣大學研究人員曾觀察到,兩隻蚯蚓並列為頭尾相對且腹面相貼的樣子,身體環帶的部位會變為扁平,肌肉會顯現出張力將對方抓牢,我們施以外力也不容易將其拉開,觀察到整個交配的過程約為 5 分鐘,最後因人為干擾過大而中斷。
從 1999 年 3 月起,台灣大學研究人員注意到參狀遠環蚓(Amynthas aspergillum)會有晚上爬出地表的行為,特別是在雨後的夜晚,此種蚯蚓長超過 30 公分,約指頭粗,即俗稱「蛇蚯蚓」的一種 ( 莊及陳,1998 ),分佈範圍從日本、大陸、香港到台灣等,在台灣廣泛分佈於平地,自台北到彰化皆有發現紀錄。參狀遠環蚓有兩對受精囊孔,分別位於 7/8-8/9 腹側的節間裂縫上,雄孔在第 18 節腹側,位於乳突圓頂,周圍有環紋,近雄孔內側會有兩大群乳突,在不同地方或不同個體間,每群乳突數量不一,約有 6-12 個。
在台北市的一些公園中,參狀遠環蚓數量還算不少,牠們會在土壤潮濕的夜間將身體前半截鑽出地表,從入夜開始,至 10 點到 12 點的數量最多,之後數量又會下降。爬出地表的參狀遠盲蚓,主要的行為多為覓食,牠們會伸出口前葉會試探周遭各個地方,當發現適當的食物會以口前葉吸住,再用力拔起或折斷該植物,除吃食土壤表層的苔蘚類外,甚至會吃下新鮮的植物葉片,我們甚至有看到此種蚯蚓將一枝長達 3 公分寬 0.5 公分的樹枝吞下。
爬出的參狀遠環蚓除了覓食外,並會進行交配,在 1999 年 5 月初曾觀察到疑似蚯蚓交配的動作,但因為干擾過大,兩隻蚯蚓迅速縮回洞中,直到 5 月 26 日,我們終於首次完整記錄到台灣蚯蚓的交配過程,其交配的行為似乎並不是特意要進行而發生,根據我們多次觀察,我們認為兩隻蚯蚓應該是在偶然遇見下進行交配:兩隻蚯蚓會在遇見後,一隻蚯蚓會先以口前葉試探對方,若另一方無交配意願,雙方會以相當快速的速度縮回洞內,否則會以頭沿著對方身體向前伸展,但動作非常緩慢,口前葉會不斷試探,然後兩隻蚯蚓各轉九十度,以利腹部相貼,當雄孔對到對方的受精囊孔時會停止再向前,這時可觀察到其雄孔突起,雄孔突出的模樣及乳突相當明顯,並且環帶部位腹部會扁平向兩邊伸展,將對方緊緊抓住,維持一段時間不動,接下來會有一隻蚯蚓開始動作,其頭部至雄孔的部位會在另一隻不動的蚯蚓受精囊孔處來回多次,不刻意停留在固定受精囊孔處,可以稍作停留約 2-5 分鐘,以便將精子傳到對方受精囊中,之後換另一隻蚯蚓進行相同的動作。國外的文獻記載中(Dubash, 1960 cited by Edward and Bohlen, 1996)環毛蚓(Pheretima)則是雄孔會先對準最後一對受精囊孔,將精子傳到受精囊中,再向前至前一對受精囊中,而這與我們觀察到的交配行為有所不同(文中未清楚描寫是否同時交換精子)。
同一晚中該對蚯蚓可能是受到人為干擾共交配了 3 次,第一次明顯受到人為干擾,在 10 分鐘後暫時分開,但過 30 分鐘左右,又再爬出互相試探重新進行交配,這次從兩隻蚯蚓互相接觸到分開為止,歷時約 1 小時,對於我們照相的閃光燈的強光毫不理會,之後兩隻蚯蚓分開,各自回到洞中。但過了 10 分鐘,一隻又爬出地表,但並未馬上向另一隻的洞穴前進,而是到處覓食,直到另一隻也爬出覓食,兩方又相遇,再進行第三次的交配,此次時間稍短,約只有 30 分鐘左右,總計自晚上 10 點到 12 點進行約 2 個小時的交配行為。在這之前, 5 月初,我們也曾發現過參狀遠環蚓的交配,但由於腳步的震動而躲回洞穴中,之後兩隻蚯蚓雖有鑽出地洞但沒有相遇,所以沒有再一次的交配行為,而是各自覓食。
從上述觀察我們認為爬出地表進行交配的參狀遠盲蚓,並未有特定交配對象,也未如某些生物會以費落蒙吸引對方來進交配,似乎是一種隨機相遇而發生的行為,會有如此的假設,主要是因為爬出的蚯蚓,不會向特定的地方爬去,而是試探似的到處覓食,直到雙方相遇才試圖交配。國外已有報告證實蚯蚓會分泌一些警示性費落蒙 ( Halpern et al., 1987;Jiang, 1989; Ressler, 1968 ),使其他蚯蚓碰觸到時產生痙攣扭動的現象,但是尚無報告顯示蚯蚓會分泌性費落蒙,吸引其他蚯蚓進行交配。
從 1999 年 3 月起,台灣大學研究人員注意到參狀遠環蚓(Amynthas aspergillum)會有晚上爬出地表的行為,特別是在雨後的夜晚,此種蚯蚓長超過 30 公分,約指頭粗,即俗稱「蛇蚯蚓」的一種 ( 莊及陳,1998 ),分佈範圍從日本、大陸、香港到台灣等,在台灣廣泛分佈於平地,自台北到彰化皆有發現紀錄。參狀遠環蚓有兩對受精囊孔,分別位於 7/8-8/9 腹側的節間裂縫上,雄孔在第 18 節腹側,位於乳突圓頂,周圍有環紋,近雄孔內側會有兩大群乳突,在不同地方或不同個體間,每群乳突數量不一,約有 6-12 個。
在台北市的一些公園中,參狀遠環蚓數量還算不少,牠們會在土壤潮濕的夜間將身體前半截鑽出地表,從入夜開始,至 10 點到 12 點的數量最多,之後數量又會下降。爬出地表的參狀遠盲蚓,主要的行為多為覓食,牠們會伸出口前葉會試探周遭各個地方,當發現適當的食物會以口前葉吸住,再用力拔起或折斷該植物,除吃食土壤表層的苔蘚類外,甚至會吃下新鮮的植物葉片,我們甚至有看到此種蚯蚓將一枝長達 3 公分寬 0.5 公分的樹枝吞下。
爬出的參狀遠環蚓除了覓食外,並會進行交配,在 1999 年 5 月初曾觀察到疑似蚯蚓交配的動作,但因為干擾過大,兩隻蚯蚓迅速縮回洞中,直到 5 月 26 日,我們終於首次完整記錄到台灣蚯蚓的交配過程,其交配的行為似乎並不是特意要進行而發生,根據我們多次觀察,我們認為兩隻蚯蚓應該是在偶然遇見下進行交配:兩隻蚯蚓會在遇見後,一隻蚯蚓會先以口前葉試探對方,若另一方無交配意願,雙方會以相當快速的速度縮回洞內,否則會以頭沿著對方身體向前伸展,但動作非常緩慢,口前葉會不斷試探,然後兩隻蚯蚓各轉九十度,以利腹部相貼,當雄孔對到對方的受精囊孔時會停止再向前,這時可觀察到其雄孔突起,雄孔突出的模樣及乳突相當明顯,並且環帶部位腹部會扁平向兩邊伸展,將對方緊緊抓住,維持一段時間不動,接下來會有一隻蚯蚓開始動作,其頭部至雄孔的部位會在另一隻不動的蚯蚓受精囊孔處來回多次,不刻意停留在固定受精囊孔處,可以稍作停留約 2-5 分鐘,以便將精子傳到對方受精囊中,之後換另一隻蚯蚓進行相同的動作。國外的文獻記載中(Dubash, 1960 cited by Edward and Bohlen, 1996)環毛蚓(Pheretima)則是雄孔會先對準最後一對受精囊孔,將精子傳到受精囊中,再向前至前一對受精囊中,而這與我們觀察到的交配行為有所不同(文中未清楚描寫是否同時交換精子)。
同一晚中該對蚯蚓可能是受到人為干擾共交配了 3 次,第一次明顯受到人為干擾,在 10 分鐘後暫時分開,但過 30 分鐘左右,又再爬出互相試探重新進行交配,這次從兩隻蚯蚓互相接觸到分開為止,歷時約 1 小時,對於我們照相的閃光燈的強光毫不理會,之後兩隻蚯蚓分開,各自回到洞中。但過了 10 分鐘,一隻又爬出地表,但並未馬上向另一隻的洞穴前進,而是到處覓食,直到另一隻也爬出覓食,兩方又相遇,再進行第三次的交配,此次時間稍短,約只有 30 分鐘左右,總計自晚上 10 點到 12 點進行約 2 個小時的交配行為。在這之前, 5 月初,我們也曾發現過參狀遠環蚓的交配,但由於腳步的震動而躲回洞穴中,之後兩隻蚯蚓雖有鑽出地洞但沒有相遇,所以沒有再一次的交配行為,而是各自覓食。
從上述觀察我們認為爬出地表進行交配的參狀遠盲蚓,並未有特定交配對象,也未如某些生物會以費落蒙吸引對方來進交配,似乎是一種隨機相遇而發生的行為,會有如此的假設,主要是因為爬出的蚯蚓,不會向特定的地方爬去,而是試探似的到處覓食,直到雙方相遇才試圖交配。國外已有報告證實蚯蚓會分泌一些警示性費落蒙 ( Halpern et al., 1987;Jiang, 1989; Ressler, 1968 ),使其他蚯蚓碰觸到時產生痙攣扭動的現象,但是尚無報告顯示蚯蚓會分泌性費落蒙,吸引其他蚯蚓進行交配。

參狀遠環蚓交配
延伸學習
